Origine de l'instabilité du génome:les éléments génitiques mobiles
On a cru longtemps que le matériel génétique était quelque chose d’immuable et de fixe, les années 40, une chercheuse du nom de Barbara Mac Clintock mettait en évidence des ries génétiques chez le maïs qu’elle ne pouvait expliquer qu’en imaginant que des parée l’ADN changeaient de place au cours du temps. La réaction fut le scepticisme général te ?). Il fallut attendre plus de 40 ans pour que ce travail soit reconnu.
On connaissait depuis longtemps la capacité des virus d’intégrer leur ADN dans l’ADN éries, ou de certains plasmides de s’intégrer dans l’ADN circulaire principal. Au début 33 70, on découvrit d’autres éléments capables de changer ainsi de place mais chromosome » bactérien et inexistants en dehors de lui, les éléments IS. On fit alors uchement avec les publications de Mac Clintock et le concept d’élément génétique fut établi. Depuis, les techniques de clonage et de séquençage du génome ont permis de nombreux éléments génétiques mobiles ou transposons.
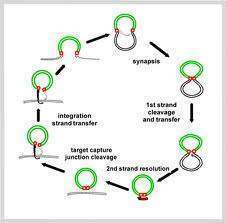
ces éléments s’avérèrent changer de place par recombinaison mais d’autres, prise générale, effectuaient de véritables sauts d’une partie à l’autre de Chacun de ces transposons contenait un certain nombre de gènes et était limité par des particulières indispensables au processus. On découvrit alors de plus en plus de tels chez les Eucaryotes. La transposition peut s’effectuer de deux façons. Soit la est simplement changée de place, soit une copie en est effectuée qui est placée h. séquence modèle restant en place. Des enzymes appelées transposases catalysent aïs, qui s’effectuent soit directement à l’état ADN soit par un intermédiaire ARN et ¿dû possède alors le gène de la transcriptase réverse, lac découverte importante fut la ressemblance entre certains de ces éléments mobiles et par certains rétrovirus qui peuvent induire des cancérisations. Alors, la cancé- serait-elle causée par des transpositions ? Les virus sont-ils des transposons échappés ou les transposons des virus intégrés dans le génome ? Ces questions sont pour résoudre les problèmes liés à l’apparition de la vie et n’ont pas reçu de ¿rSiutives pour l’instant.
emplacement de ces gènes n’est pas un événement sans conséquence dans la mesure des gènes voisins peut en être modifiée. D’autre part, ils peuvent, en s’inté- ~ une insertion ou une délétion locale ou tout au moins perturber la transcrip- deux éléments identiques sont présents sur un chromosome ils peuvent se transposer ensemble, emportant avec eux la séquence comprise entre eux, formant transposon complexe.
Transposition et variation
On connaît chez la levure Saccharomyces cerevisiae deux types sexuels a et a cellules haploïdes. Lors d’un croisement, les deux cellules sont obligatoirement de types rents, mais le type peut changer dans la descendance, d’une génération à l’autre. 0« maintenant que ce caractère est sous la dépendance d’un locus unique MAT bi (Watson et al. 1994), mais celui-ci est «entouré» par une copie supplémentaire de u des deux allèles a et a. Chacune de ces copies peut s’apparier successivement avec le MAT et y provoquer la transformation de ce dernier . Le mécanisme mol ‘ est très semblable à la transposition des éléments Tn3 des Procaryotes. L’intérêt pour Iules à haute fréquence de division est vraisemblablement de permettre la réalisation « brassage » maximum dans la population c’est-à-dire d’étendre les possibilités de naison.
Le mécanisme de la variation des antigènes de surface chez le trypanosome est fait comparable. La fréquence des transpositions réplicatives y est cependant moins élev la banque de séquences alternatives est beaucoup plus importante. Les séquences non e mées sont déplacées vers des sites d’expression associés aux télomères, les extrémités chromosomes. Ce déplacement laisse en place une copie de la séquence déplacée et d généralement, la séquence qui occupait le site d’expression. Ce changement périodique antigènes de surface permet ainsi au parasite d’échapper au système immunitaire de l’hôte.
Enfin, le cas des immunoglobulines est un dernier exemple d’une sélection par génomes des propriétés de transposition de certaines séquences. La contrepartie est que transpositions se font parfois de telle sorte qu’elles perturbent certains mécanismes fc mentaux de la régulation de l’expression de l’information génétique. C’est le cas des lyn mes, les cancers des lymphocytes, où des proto-oncogènes normalement non exprimes retrouvent après transposition à proximité de séquences induisant leur expression. Pour Watson et al. (1994) il s’agit du « prix à payer pour avoir développé une machinerie complexe de transposition d’ADN pour élaborer notre système immunitaire ».
Transposition et inflation du génome
De très nombreux autres éléments génétiques mobiles semblent n’avoir pour toute fonction que d’assurer leur propre survie. Les éléments IS, ou séquences d’insertion, des Proca- ryotes par exemple se déplacent d’un endroit à l’autre du chromosome. Il en est de même pour ies éléments Ac-Ds du maïs, responsables du panachage des épis étudié par B. Mac Clintock. Ces transposons peuvent représenter près de 10% du génome d’une cellule et bien que la probabilité individuelle de transposition de chaque séquence soit faible, leur nombre fait que la isposition est fréquente. On estime par exemple que près de la moitié des mutations spontanées étudiées chez la drosophile sont dues à des insertions de transposons. Ces éléments se déplacent de façon conservative ou réplicative mais provoquent toujours la duplication d’une courte séquence du site d’insertion. L’accumulation d’événements de transposition ne peut alors qu’aboutir à l’augmentation continuelle de la quantité d’ADN présente dans un génome.
En dehors de ces courtes duplications induites aux sites d’insertion des éléments génétique mobiles, la formation de transposons complexes avec transposition réplicative est sans te un puissant mécanisme pour la réalisation de duplications géniques .
L’augmentation de la quantité d’ADN peut également se révéler sous d’autres formes, séquences répétées un très grand nombre de fois, satellites et microsatellites, ou des séquences inactives gardant la trace d’une activité passée. Dans la première catégorie les séquences Alu, répétées plusieurs centaines de milliers de fois dans un génome ain, dont la structure apparaît proche de celle des transposons en particulier du fait de la ence de deux séquences répétées et inversées aux extrémités. Dans la seconde se placent pseudogènes maturés dont la structure paraît directement dérivée de celle d’un ARN mes- après excision et épissage des introns. Dans le cas du pseudogène de la P-tubuline aine, on retrouve très exactement cette situation .
L’absence apparente de fonction bien précise de ces différentes séquences a conduit les ‘heurs à les qualifier d’ADN égoïste. Cependant, la transposition peut être considérée e un moyen d’augmenter le nombre de combinaisons de gènes et donc la diversité génie. Elle permettrait de « tester » des effets de position, retenus ou éliminés par la sélection elle. On a pu montrer en effet chez la drosophile que certains transposons sont systéma- îent associés à des remaniements chromosomiques du type inversion ou translocation, transposition permet également d’activer ou d’inactiver certains gènes, dans la mesure où ins rétrotransposons possèdent des séquences promotrices capables de déclencher la -cription des séquences situées en aval de leur point d’insertion.
On le voit, la transposition apparaît de plus en plus comme un mécanisme majeur de l’évolution.
Vidéo : Origine de l’instabilité du génome:les éléments génitiques mobiles
Vidéo démonstrative pour tout savoir sur : Origine de l’instabilité du génome:les éléments génitiques mobiles
